谷禾健康

以前我们科普过肠道菌群在门级别水平分类的肠道细菌四大常见菌门——拟杆菌门,厚壁菌门,变形菌门,放线菌门。
详见:肠道细菌四大“门派”——拟杆菌门,厚壁菌门,变形菌门,放线菌门
但是随着研究范围以及样本的扩大,发现我们人体肠道内除了这些常见的菌群之外,还有许多小众门派的菌群在肠道平衡中也扮演着重要的角色,它们同样可以帮助我们消化食物、维持肠道健康、增强免疫力等等。这些微生物组通常作为宿主基因组的功能扩展,在宿主生理和新陈代谢的调节中起着至关重要的作用。
在本文中,我们将深入探讨这些小众门派的菌群,揭示它们的神秘力量和重要性,帮助我们更好地了解肠道菌群的多样性和复杂性。
我们根据谷禾肠道数据库人群检出比例的丰度排名依次介绍这些菌及其与肠道和人体健康的相关信息。
根据人群检出丰度依次为:
疣微菌门 Verrucomicrobia
梭杆菌门 Fusobacteria
蓝藻门 Cyanobacteria
酸杆菌门 Acidobacteria
软壁菌门 Tenericutes
绿弯菌门 Chloroflexi
互养菌门 Synergistetes
芽单胞菌门 Gemmatimonadetes
黏胶球形菌门 Lentisphaerae
浮霉菌门 Planctomycetes
硝化螺旋菌门 Nitrospirae
脱铁杆菌门 Deferribacteres
螺旋体 Spirochaetes
装甲菌门 Armatimonadetes
绿菌门 Chlorobi
迷踪菌门 Elusimicrobia
衣原体 Chlamydiae
这里我们对门层级中的常见肠道菌群逐个介绍。
No.01
疣 微 菌 门 Verrucomicrobia
疣微菌门(Verrucomicrobia)是一类革兰氏阴性细菌,是细菌分类学中的一个门级分类单元。这类细菌的细胞形态多样,包括球形、杆状、螺旋形等,常见于土壤、水体和动物肠道等环境中。
疣微菌门是细菌域内的一个新分支,1997年被列为一个门。它们代表系统发育树中的一个独特谱系,包含许多环境物种以及少量培养物种。
疣微菌门是一类广泛存在于自然环境中的细菌,包括多种典型的菌种,例如常听说的Akkermansia muciniphila,其他还有Opitutus terrae、Prosthecobacter debontii等。这些菌种在不同的环境中具有不同的代谢和生理特性。
▼
代谢特性
疣微菌门存在于肠道粘膜内层,在健康个体中大量存在,它们可以分解多糖类物质,如黏多糖和纤维素等,从而提供能量和营养物质。
疣微菌门还可以产生短链脂肪酸,如丙酸和丁酸等,这些物质对肠道健康和免疫系统的调节具有重要作用。
虽然说疣微菌门仅占肠道微生物群落总数的一小部分,但结果表明,一些疣微菌种系型对多糖和木聚糖的降解做出了重大贡献。基因组编码多种糖苷水解酶、硫酸酯酶、肽酶、碳水化合物裂解酶和酯酶,具有水解多种多糖的机制。
▼
健康相关
在肠道微生物组中,疣微菌门是一个重要的菌群之一。 谷禾肠道数据库中大约56.29%的人群有检出。
⇒
研究表明,疣微菌门的丰度与肠道健康密切相关。疣微菌门有助于人体肠道的葡萄糖稳态, 具有抗炎特性,可进一步帮助肠道健康。研究表明 该菌与foxp3 基因之间存在正相关关系,foxp3 基因是一种在人类中表达抗炎和免疫力的基因。
⇒
一些疾病如肥胖症、炎症性肠病、睡眠障碍和2型糖尿病等,与疣微菌门的丰度降低有关。其他如哮喘、自闭症等疾病人群中丰度也会变化。
⇒
其中,Akkermansia muciniphila(简称AKK菌)是疣微菌门中研究较多的菌种之一。研究表明,Akk菌可以降低肥胖、糖尿病、肠炎、肠癌等疾病的风险。这是因为Akk菌可以促进肠道黏液层的生长和维护,增强肠道屏障功能,减少有害菌的生长,降低肠道内毒素的水平,从而保护肠道健康。
详见:
Nature | AKK菌——下一代有益菌
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
No.02
梭 杆 菌 门 Fusobacteria
梭杆菌门 (Fusobacteria)是细菌门之一,它们是一类革兰氏阴性菌,通常是长杆状或螺旋状的。在谷禾肠道数据中,该菌的检出率是49.16%.
梭杆菌门包括多种典型的种属,如:Fusobacterium nucleatum(具核梭杆菌)、
Fusobacterium varium(变异梭杆菌)、
Fusobacterium necrophorum等。
▼
生存环境
梭杆菌门广泛存在于自然环境中,包括土壤、水体和动物肠道等。已证明梭杆菌门下的许多物种可以自由生活,而无需与周围环境中的其他生物结合。其中大部分包括在海洋环境中发现的物种,例如 Llyobacter 属和 Psychrilyobacter 属的成员。
Photograph by Hipersynteza
自由生活的梭杆菌属的最好例子之一是 Psychrilyobacter atlanticus 物种,它可以在大西洋的海洋沉积物中找到。
它们是一类厌氧菌,通常生长在肠道内的低氧环境中。梭杆菌门在人体内的分布广泛,不仅存在于肠道中,还存在于口腔、阴道和皮肤等部位。
▼
健康相关
⇒
一些研究表明,梭杆菌门可能与肠道疾病有关。例如,一些研究发现,梭杆菌门在结肠癌组织中的含量明显高于正常组织。此外,梭杆菌门也与炎症性肠病(IBD)和肠道感染有关。
⇒
在口腔中,梭杆菌门的存在则与龋齿和牙周炎等口腔疾病有关。
然而,梭杆菌门在肠道菌群中也扮演着有益的角色。梭杆菌门可以产生一些对人体有益的代谢产物,如丙酮酸和丁酸等。一些研究表明,梭杆菌门可能参与了肠道菌群的稳态维持和代谢功能。
⇒
吸烟者的梭杆菌丰度显著更高(P = 0.009,FDR = 0.027 )。
⇒
每日食用面包的人群中,梭杆菌丰度也显着更高(P = 0.005,FDR = 0.015)。
⇒
饮用更多咖啡的个体表现出略显着更高的梭杆菌属丰度( P= 0.02,FDR = 0.20)。
总的来说,梭杆菌门在人体健康中的作用还需要进一步的研究和探索。虽然它们可能与一些疾病有关,但在肠道菌群中的作用也不容忽视。
扩展阅读:梭杆菌属Fusobacterium——共生菌、机会致病菌、致癌菌
No.03
蓝 细 菌 门 Cyanobacteria
蓝细菌门(Cyanobacteria),是一类原核生物,也被称为蓝藻或蓝藻菌门。
蓝绿藻植物菌门下物种又称蓝细菌、蓝绿菌、蓝藻或蓝菌,包括蓝鼓藻、蓝球藻等生物。过去曾长期被归于藻类,但实际上蓝菌与真核生物非常不同,例如没有核膜,没有细胞器,其遗传物质DNA也不构成染色体,这些都是细菌的特征,因此现时已被归入细菌域。
▼
生存环境
蓝细菌分布极广,普遍生长在淡水、海水和土壤中,主要分布在含有机质较多的淡水中;在极端环境(如温泉、盐湖、贫瘠的土壤、岩石表面或风化壳中、冰雪上、植物树干等)中也能生长;蓝菌有些还可穿入钙质岩石(如钙藻类)或土壤深层中(如土壤蓝藻),故有“先锋生物”的美称。
▼
代谢特性
蓝菌是一类能透过产氧光合作用获取能量的细菌,但有些也能透过异营来获取能量。
蓝绿菌在地球上已存在约21亿年,是目前以来发现到的最早的光合放氧生物,对地球表面从无氧的大气环境变为有氧环境起了巨大的作用。通过刺激生物多样性和导致厌氧生物接近灭绝,显著的改变了在地球上生命形式的组成。根据内共生学说,在植物和真核藻类发现的叶绿体是从蓝细菌祖先通过内共生进化而来的。
▼
常见菌
以下是一些常见的典型菌种:
Anabaena(鱼腥藻):这是一种常见的蓝藻,可以在淡水和海水中生长。它们通常形成长链,其中一些细胞可以进行氮固定,这对于生态系统的氮循环非常重要。
Microcystis(微囊藻):这是一种广泛分布的蓝藻,可以在淡水和海水中生长。它们通常形成大量的胞囊,这些胞囊可以释放出毒素,对水生生物和人类健康造成威胁。
Spirulina(螺旋藻):这是一种广泛应用于食品和保健品的蓝藻。它们富含蛋白质、维生素和矿物质,被认为具有多种健康益处。
▼
健康相关
谷禾肠道数据库中大约39.06%的人群有检出,但一些研究表明,它们可能与肠道健康和一些疾病有关。
一些研究发现,蓝细菌在肠道中的丰度与炎症性肠病(如克罗恩病和溃疡性结肠炎)的发生有关。
蓝细菌中的一些代表性菌种,如前面提到的Anabaena sp.和Microcystis sp.,可能产生毒素,对人体健康造成威胁。
⇒
在一些污染的地区,其中绝大多数淡水湖泊中发现了大量的蓝藻水华。蓝藻毒素早就被认为通过肝脏影响与之相关的健康问题,食用受蓝藻污染的饮用水相关的健康问题也包括胃肠炎和肠道不适,其中肠道微生物组发挥了关键作用。但蓝藻毒素的作用机制仍然是一个关键的、大部分未被探索的问题。
肠道蓝细菌丰度与人类健康和疾病的潜在联系

doi.org/10.3390/life12040476
⇒
在谷禾肠道样本人群中,发现渐冻症部分群体里发现蓝藻门(蓝细菌)比较高。结合研究发现蓝菌门生物皆含有神经毒素BMAA(β-N-methylamino-L-alanine),并可能透过食物链不断累积产生生物放大作用,对人类的损害可能会逐渐增加。BMAA已证实会对动物产生强烈的毒性,加速动物脑神经退化、四肢肌肉萎缩等等,小量BMAA积累对小鼠已能选择性杀死神经元。
因此需要注意,在水体中的过度生长会导致水体富营养化,产生毒素,对人体健康和生态环境造成危害。需要加强对蓝细菌的监测和管理。
No.04
酸 杆 菌 门 Acidobacteria
酸杆菌门 (Acidobacteria) ,虽然这个名字听起来都是嗜酸性的,但一些物种可以在中性和弱碱性环境中找到。
虽然一些研究表明酸杆菌门有超过 18 个类别,但只有三个类别是众所周知的。它们是:
- Acidobacteriia
- Blastocatellia
- Holophagae
(Holophagae 纲在某些文献中被描述为一个目)
▼
常见菌属
下面了解一下酸杆菌门下的3个常见属:
◗ 酸杆菌属
酸杆菌门下第一类重要的菌属——酸杆菌属(Acidobacterium Genus),通常存在于酸性环境中。
荚膜酸杆菌(Acidobacterium capsulatum)是该组中最受欢迎的成员之一(由大约 8 个菌株组成)。与该类别的其他成员一样,荚膜酸杆菌是一种革兰氏阴性细菌。
酸杆菌属也是好氧菌,它们本质上是嗜温的;在适中的温度(20~45°C之间)生长良好。
与这些细菌相关的一些其他特征包括:
- 不形成/产生孢子
- 大部分品种呈杆状(拉长)
- 它们分解糖以获取能量(它们是糖分解的)——一些用作能量来源的糖包括葡萄糖、麦芽糖、纤维二糖和淀粉等。
- 通过鞭毛(peritrichous flagella)移动
- 栖息地 pH 值范围在 3.0-6.0 之间的
◗ Terracidiphilus属
酸杆菌门下第二类重要的菌属——Terracidiphilus 属,由已知可产生用于分解几丁质和寡糖的细胞外酶的生物体组成。
该属中最受欢迎的物种之一是 Terracidiphilus gabretensis。这种细菌常见于针叶林中,它在碳汇中起着重要作用。
- 革兰氏阴性细菌
- 杆状(或卵形)
- 通过鞭毛移动并且不产生孢子
- 只生长在有氧环境中
- 作为一种嗜酸性生物,它可以在 pH 值在 3 到 6 之间的酸性环境中找到
- 它也是一种嗜温细菌,最佳生长温度在 20 到 24 °C 之间。
◗ Terriglobus 属
Acidobacteria 门下第三类重要的菌属——Terriglobus属,是革兰氏阴性菌,与 Granulicella 和 Adaphobacter 属密切相关。
该组的成员是土壤中常见的好氧化学有机异养生物。虽然这些生物通常存在于土壤(根际土壤)中,但在淡水生境中也有。
目前,该属有 5 个知名物种,其中包括:
- T. roseus
- T. tenax
- T. albidus
- T. saanensis
- T. aquaticus
该属的一些特征为:
- Terriglobus 细菌呈弱酸性,在 pH 值介于 5 和 6 之间时生长最佳。
- 也是嗜温性的,喜欢 25-30℃ 之间的温度。
- Terriglobus 的形成可能具有粉红色色素沉着的圆形菌落。
- 除了碳源(提供能量的糖)之外,它们还需要氮来维持生存(从氨基酸和氯化铵中获取)。
图源:Biology LibreTexts
与这些细菌相关的一些其他特征包括:
- 不活动:它们不像其他一些酸杆菌那样使用鞭毛移动
- 长度从 0.8-3um,直径从 0.4-0.9um不等
▼
代谢特性
据估计,酸杆菌是土壤中主要菌群,约占土壤中所有微生物的 20%。
在环境中,酸杆菌在养分循环中起着重要作用。
碳循环 ——通过降解各种碳源(糖和蛋白质等),这些细菌将碳返回到环境中。然后,这些碳被植物和其他生物用于各种功能,循环继续。
氮循环 ——目前,Geothrix fermantans是酸杆菌门内唯一已知在氮循环中发挥一定作用的物种。通过减少硝酸盐来实现的。
硫循环 ——嗜热氯酸杆菌是需要硫来生长和发育的酸杆菌的一个例子。硫代谢有助于硫循环。
▼
健康相关
与人体肠道和健康的关系方面,目前对于酸杆菌的研究还比较有限。
一些研究表明,酸杆菌门可能在人体肠道中存在,谷禾肠道数据库中大约33.05%的人群有检出。
肠道微生物群落的失衡与多种疾病的发生有关,而酸杆菌门的数量在某些疾病中可能会发生变化。
⇒
一些研究表明,肠道炎症性疾病患者的肠道中酸杆菌门的数量较低。
⇒
妊娠期糖尿病患者中酸杆菌门显著增加,并与血糖水平呈正相关。
⇒
在特发性肾病综合征患者中,酸杆菌门显著减少。
⇒
糖尿病肾病具有与健康对照不同的肠道微生物群,酸杆菌门在糖尿病肾病患者中增加。
扩展阅读:2型糖尿病如何做到可防可控?肠道菌群发挥重要作用
⇒
一项研究中,幽门螺杆菌阳性受试者的微生物群落表明变形杆菌、酸杆菌和螺旋体的数量增加。
扩展阅读:正确认识幽门螺杆菌
但是,目前还需要更多的研究来探究酸杆菌门与人体肠道和健康之间的具体关系。
No.05
软 壁 菌 门 Tenericutes
软壁菌门(Tenericutes),在谷禾肠道数据中,该菌的检出率是29.61%.
这些细菌通常是无细胞壁的,因此它们的形态非常多样化,也被称为无壁菌门。由质膜包围的细胞组成的革兰氏阴性菌。
软壁菌门由从厚壁菌门进化而来的细菌组成。尽管一些研究人员强烈认为软壁菌门应该被整合到厚壁菌门中,但两个显着特征使软壁菌门有别于厚壁菌门:
- 无法合成肽聚糖的前体,从而形成细胞壁
- 基因组大小的大大减小(在530-2220kbp之间)
目前,软壁菌门的分类地位尚不确定。随着未来鉴定出更多新的软壁菌门细菌谱系,软壁菌门的分类定位和单系性可能会受到进一步挑战。
▼
生存环境
软壁菌门普遍存在于许多环境中。16S rRNA 测序已经在包括深海在内的不同环境中识别出大量未知的软壁菌门进化枝,这表明这些细菌可能代表独立生活的微生物,其生活方式与宿主无关。
事实上,在深海冷泉和盐水池中分别发现了Candidatus Izemoplasma和 Haloplasma自由生活。这些深海自由生活的软壁菌门细菌表现出新陈代谢的多样性和适应性的灵活性,表明有可能从海洋甚至其他极端环境中分离出更多的软壁菌门细菌。
▼
健康相关
⇒
一项关于老年2型糖尿病认知障碍的研究发现参与者中,认知障碍患者的血红蛋白和高密度脂蛋白水平较低,相对糖尿病对照组而言,认知障碍糖尿病患者的软壁菌门Tenericutes丰度较低。
⇒
在长寿村社区老年人的肠道菌群中,厚壁菌门、软壁菌门Tenericutes和放线菌门的相对丰度显著低于城市化城镇社区。
扩展阅读:健康长寿的步伐永不停歇
⇒
一项研究将86名儿童(5-15岁)被分为三组:
- 代谢健康肥胖(n = 42)
- 代谢不健康肥胖(n = 23)
- 健康的正常体重对照组(n = 21)
与对照组相比,在代谢性不健康肥胖受试者中,软壁菌门(Tenericutes)以及α和β多样性显著降低。与对照组相比,互养菌门(Synergistetes)和拟杆菌属在代谢健康肥胖人群中更为普遍。
总的来说,软壁菌门在肠道中的作用和其与健康的关系还需要进一步的研究和探索。
注意,软壁菌门在临床上比较难分辨。它们往往不太可能生长,也不太可能被经典微生物学技术识别,一般通常需要进行分子鉴定。属内的敏感性特征通常变化很大,这使得针对他们的特异性鉴定以及合理选择抗菌药物非常重要。
No.06
绿 弯 菌 门 Chloroflexi
绿弯菌门(Chloroflexi)是一类光合细菌,可以利用光合作用产生能量。又称作绿非硫细菌,还有一部分称作热微菌的细菌也属于绿非硫细菌。绿弯菌门的细菌生活在海洋,淡水等环境中。
该门包括六类:
- Chloroflexi
- Anaerolineae
- Caldilineae
- Ktedonobacteria
- Dehalococcoidetdia
- Thermomicrobia
▼
代谢特性
绿弯菌门由不同的生物群组成,包括无氧光合自养生物、好氧化学异养生物、嗜热生物以及通过有机氯化化合物的还原脱卤获得能量的厌氧生物。
典型的绿弯菌门细菌是线形的,通过滑行来移动。它们是兼性厌氧生物,在光合作用中不产生氧气,不能固氮。利用3-羟基丙酸途径,而不是常见的卡尔文途径来固定二氧化碳。
图源:de-academic
所有已知的成员都是丝状的,具有不寻常的滑动机制作为一种运动方式,虽然大多数革兰氏染色呈阴性,但没有一个具有革兰氏阴性菌特有的脂多糖外膜。
绿弯菌门包含生态和生理上多样化的细菌群,已在越来越广泛的厌氧生境中检测到这些细菌,包括沉积物、温泉、产甲烷厌氧污泥消化池,它们在这些地方非常丰富,并发挥着重要的发酵作用有助于污泥粒化。绿弯菌门是固体废物和废水处理系统中最主要的门之一。特别是,Anaerolineae 类已被确定为全面厌氧反应器中的核心微生物种群之一。
绿弯菌门可能参与了肠道中的一些代谢过程,例如氨基酸、葡萄糖和脂肪酸的代谢。
▼
健康相关
虽然绿弯菌门在人群中不是常见菌,但目前人体肠道和口腔中也逐步检测到绿弯菌门细菌,谷禾肠道数据库中大约25.28%的人群有检出。
⇒
在门水平上,利福昔明治疗后腹泻型肠易激综合征患者的绿弯菌门(Chloroflexi)(P=0.008)、Deinococcus-Thermus菌(P=0.038)和酸杆菌群(P=0.028)增加。
⇒
在门水平上,与格雷夫斯病相比,格雷夫斯眼眶病患者中Deinococcus-Thermus菌和 绿弯菌门(Chloroflexi) 的比例显着降低。
⇒
小型研究发现,新冠肺炎刚痊愈的人与健康对照相比,绿弯菌门(Chloroflexi)显著降低。
扩展阅读:阳康后是否会二次感染,长新冠与肠道菌群的关联,多种潜在的相关干预措施
此外,一些研究还发现,绿弯菌门的存在与一些肠道疾病,如炎症性肠病和肠道肿瘤等有关联,但具体的机制还需要进一步研究。
No.07
互 养 菌 门 Synergistetes
互养菌门(一般翻译为 Synergistetes),也有翻译为增效菌门或协同菌门。在谷禾肠道数据中,该菌的检出率是24.61%.
Synergistetes细菌是最近认识到的一个门,其中已分离出 40 种生物,并且有超过三百个 16S rRNA 序列可用。
这个门的分类学历史很短,最近才被确定为细菌域内的一个独立门。第一个代表性物种,Synergistes jonesii,最初从夏威夷山羊的瘤胃中分离出来,以其命名,最初被分类在 Deferribacteres 门中。
▼
代谢特性
来自该门的物种共有的表型特征包括它们的革兰氏阴性细胞壁、厌氧、杆状/弧菌状细胞形状
图源:researchgate
虽然脂多糖存在于双层细胞膜中是一个重要特征,但在互养菌门物种中尚未被报道,但它们确实含有参与脂多糖生物合成的各种蛋白质的基因。虽然有些物种不能分解糖,但所有互养菌门都具有发酵氨基酸的能力。
它们可以利用多种有机物作为碳源和能源。
一些Synergistetes菌属可以利用蛋白质、脂肪酸和多糖等有机物进行代谢,同时还可以参与肠道中的硫循环和氮循环等过程。
一些Synergistetes菌属还可以产生一些对人体有益的代谢产物,如短链脂肪酸等。
▼
生存环境
互养菌门主要栖息在厌氧环境中,包括动物胃肠道、土壤、油井和废水处理厂,它们也存在于人类疾病部位,如囊肿、脓肿和牙周病区域。
由于它们存在于疾病相关部位,互养菌门被认为是机会性病原体,但它们也可以在健康个体的脐部微生物组和正常阴道菌群中发现。
该门的其他物种已被确定为厌氧消化池中用于生产沼气的污泥降解的重要贡献者,并且是通过生产氢气用于可再生能源生产的潜在候选者。
常见菌属:Aminiphilus是一类革兰氏阴性菌,通常生长在富含有机物的水体中。它们可以利用氨基酸和蛋白质等有机物作为碳源和能源。
▼
健康相关
⇒
大多数人类培养的菌株来自感染部位,表明互养菌门在感染过程中增殖。互养菌门的不同分支和物种表现出不同的感染倾向:
- Cloacibacillus spp. 通常在腹膜液样本中
- J.anthropi在软组织感染中发现
- 口腔主要受到F.fastidiosum和P.piscolens感染的影响
⇒
在牙周炎患者中发现了一些互养菌门OTU,并且在牙周炎患者的患病部位龈下菌斑中比健康部位更丰富。
⇒
2型糖尿病和牙周炎患者的微生物群比非糖尿病牙周炎患者的微生物群显示出更少的互养菌门。
⇒
在坏死性溃疡性牙龈炎的病例中,互养菌门聚类A OTUs的检测水平和比例高于牙龈炎期间。
⇒
研究显示,精神病患者患抗精神病药引起的便秘的风险很高,互养菌门在便秘组的肠道微生物群中显著增高。
⇒
互养菌门可能保护甲状腺,在格雷夫斯病患者的肠道菌群中,互养菌门与促甲状腺激素受体抗体(TRAb)水平呈负相关。
No.08
芽 单 胞 菌 门 Gemmatimonadetes
芽单胞菌门(Gemmatimonadetes)是一类革兰氏阴性菌,包括:
芽单胞菌纲(Gemmatimonadetes)
芽单胞菌目(Gemmatimonadales)
芽单胞菌科(Gemmatimonadaceae)
芽单胞菌门目前仅有一属得到正式命名,即芽单胞菌属(Gemmatimonas),是一类革兰氏阴性细菌,通过出芽方式繁殖。
图源:alchetron
芽单胞菌门可以在各种环境中生长,包括土壤、淡水、海水和沉积物等。在肠道中,芽单胞菌门也是一种常见的微生物群落成员。
▼
代谢特性
芽单胞菌门可以利用多种有机物和无机物作为碳源和能源,包括葡萄糖、氨、硝酸盐、硫酸盐等。此外,芽单胞菌门还可以在低氧或缺氧条件下生长,并且能够耐受一定的重金属和有机污染物。
一些研究表明,芽单胞菌门可能在土壤和水体中发挥重要的生态功能,如有机物分解和氮循环等。此外,芽单胞菌门可能与一些环境污染物的降解有关。因此,芽单胞菌门在环境保护和生态平衡方面具有重要的作用。
芽单胞菌门的代表性菌种包括:
Gemmatimonas aurantiaca、
Gemmatimonas phototrophica等。
这些菌种具有一些特殊的代谢特征,例如Gemmatimonas aurantiaca可以利用多种有机物作为碳源和能源,同时还具有一定的光合作用能力。
▼
健康相关
芽单胞菌门与人体健康的关系尚未得到充分研究。谷禾肠道数据库中大约20.11%的人群检出。
一些研究发现,芽单胞菌门可能与肠道炎症的发生有关。
⇒
可能与骨质疏松相关:
芽单胞菌门和绿弯菌门,在原发性骨质疏松症患者和 正常对照组,以及骨质减少患者和正常对照组之间存在显着差异 ( p < 0.01 ) 。
⇒
补充唾液酸Neu5Ac对小鼠肠道形态、肝功能和肠道微生物影响的研究表明,肠道微生物群组成呈剂量依赖性变化,在门水平上,芽单胞菌门显着增加。
注:N-乙酰神经氨酸 (Neu5Ac) 是人类唾液酸的主要形式。
需要进一步的研究来探究芽单胞菌门与人体健康的关系。
No.09
黏 胶 球 形 菌 门 Lentisphaerae
黏胶球形菌门(Lentisphaerae)是一个相对较小的门,球形或椭圆形,通常是厌氧或微好氧的,可以生长在不同的环境中,如土壤、淡水、海水、动物肠道等。在谷禾肠道数据中,该菌的检出率是18.42%.
▼
代谢特性
黏胶球形菌门的代谢特征是多样的,包括无氧呼吸、发酵、光合作用等。
一些菌属可以利用多种碳源和氮源进行生长,如Lentisphaera和Victivallis可以利用多种碳水化合物和蛋白质作为碳源和氮源。
一些Lentisphaerae菌属还具有产生酸和气体的能力,如Fibrobacteres可以产生乳酸和乙酸。
Lentisphaerae还被发现可以参与到肠道中的多糖代谢和蛋白质降解等代谢过程中。
在黏胶球形菌门内的Victavallales属,可以酶解唾液酸、岩藻糖、半乳糖和 N-乙酰氨基葡萄糖,降解粘蛋白,其糖基水解酶图谱与AKK菌非常相似。
▼
健康相关
⇒
一项研究发现,黏胶球形菌门(Lentisphaerae)在健康人的肠道中的丰度较高,而在患有炎症性肠病和非酒精性脂肪性肝病 (NAFLD) 的患者中的丰度较低。
⇒
研究还发现更好的睡眠质量与更好的认知灵活性和更高比例的肠道微生物门Verrucomicrobia和Lentisphaerae有关。
⇒
也有研究发现黏胶球形菌门( Lentisphaerae)与帕金森疾病相关。
扩展阅读:肠道微生物与帕金森以及相关影响因素
⇒
可能与他汀类药物的代谢有关,高脂血症患者接受了 10 mg/天的瑞舒伐他汀治疗 4-8 周,蓝细菌门和黏胶球形菌门(Lentisphaerae)与低密度脂蛋白胆固醇水平呈正相关。
⇒
横断面研究发现,黏胶球形菌门(Lentisphaerae)在多发性硬化患者中显著降低。
扩展阅读:肠道微生物群在多发性硬化中的作用
⇒
经常食用面包的受试者的肠道微生物群中,互养菌门和黏胶球形菌门(Lentisphaerae)的相对丰度显着更高(分别为P = 0.009,FDR = 0.028 和P = 0.004,FDR = 0.011)。
No.10
浮 霉 菌 门 Planctomycetes
浮霉菌门(Planctomycetes)形成了一个独特的细菌门,具有独特的特征组合,例如,缺乏肽聚糖的蛋白质细胞壁,以及在细胞质内形成独立隔室的细胞内膜。
浮霉菌门的细胞结构
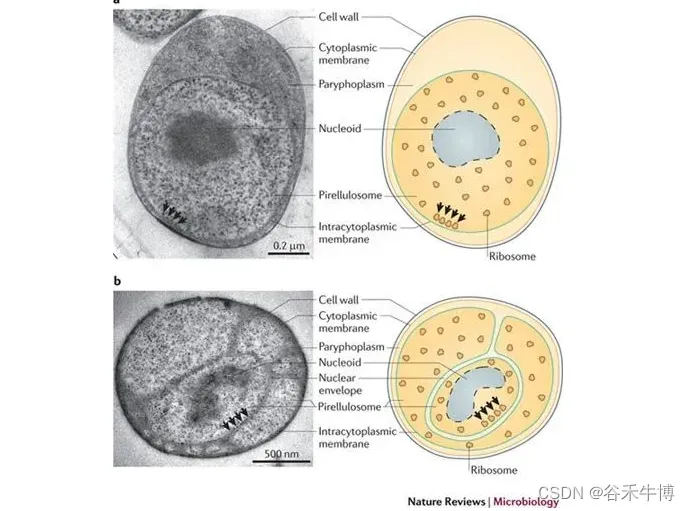
doi: 10.1038/nrmicro2578.
▼
生存环境
浮霉菌门包括多个典型菌属,如Planctomyces、Gemmata、Pirellula、Rhodopirellula等。这些菌属通常是好氧或微好氧的,可以生长在不同的环境中,如海水、淡水、土壤、沉积物、动物肠道等。主要与大型藻类、海绵和地衣等颗粒或生物相关,具体取决于物种及其硫酸酯酶可代谢的多糖。大多数浮霉菌门生长在 pH 值从 3.4 到 11 不等的营养贫乏的贫营养环境中。
大多数浮霉菌门是嗜温的,但有一些 浮霉菌门是嗜热的(50°C -60°C)。通常添加的营养素有 N-乙酰氨基葡萄糖、酵母提取物、蛋白胨和一些微量元素和大量元素。
▼
代谢特性
一些浮霉菌属还具有产生酸和气体的能力,如Pirellula可以产生乳酸和乙酸。另外,浮霉菌门还具有一些特殊的代谢特征,如一些菌属可以利用甲烷和硫化氢作为能源和碳源,如Anammoxoglobus和Brocadia。
浮霉菌门包括一些具有非常不寻常的生理学的物种,比如一些浮霉菌可以合成甾醇,这是一种真核生物的典型能力,在细菌中并不常见。
浮霉菌门的另一个不寻常的代谢特征是它们拥有编码C1转移酶的基因。这些酶以前只在产生甲烷的古细菌和一组甲烷氧化的变形菌中被发现,它们在具有一个碳原子的化合物的代谢中发挥作用。比较基因组学和蛋白质组学表明,厌氧氨氧化和非厌氧氨氧化浮霉菌之间的区别超出了铵的代谢。
▼
健康相关
浮霉菌门是人类消化道微生物群的一部分。谷禾肠道数据库中大约13.92%的人群有检出。它们的多样性因环境而异,包括个体的地理起源和抗生素治疗。
⇒
在两名患有白血病和再生障碍性中性粒细胞减少症的发热患者的血液中检测到与浮霉菌门密切相关的 DNA 序列。
⇒
G. massiliana的分离来自靠近这些患者的医院供水系统,可能支持消化道进入途径的假设,即摄入受污染的水然后在免疫功能低下的患者的血液中易位。Gemmata属的可能作为潜在的机会性病原体进入消化道。
No.11
硝 化 螺 旋 菌 门 Nitrospirae
硝化螺旋菌(Nitrospirae) 是革兰氏阴性菌,通常呈螺旋状。以其氧化亚硝酸盐和硝酸盐的能力而闻名。谷禾肠道数据库中大约11.04%的人群有检出。
硝化螺旋菌存在于各种环境中,包括土壤、淡水、海洋栖息地、污水处理厂等。在亚硝酸盐和硝酸盐等含氮化合物含量高的环境中,它们尤其丰富。
▼
代谢特性
硝化螺旋菌作为一种好氧化学自养亚硝酸盐氧化细菌,在硝化过程中发挥着关键作用。这些细菌通常与氨氧化细菌或古菌密切相关,这些细菌将氨转化为亚硝酸盐,亚硝酸盐被硝化螺旋菌进一步氧化为硝酸盐。
然而,在“相互喂食”的相互作用中,硝化螺菌也可以用尿素或氰酸盐释放的氨提供氨氧化剂,尿素或氰酸酯被进一步硝化。
图源:Mmolecular
硝化螺旋体成员甚至单独催化两个硝化步骤,因此被称为完全氨氧化剂或“comammox”生物体。这与传统的硝化细菌不同,传统的硝化细菌需要两种不同类型的细菌来完成这些步骤。
一些硝化螺旋菌菌株利用H2和甲酸盐等替代底物,使用氧气或硝酸盐作为末端电子受体,并可以在好氧亚硝酸盐氧化的同时利用这些能源。这种代谢的多样性使硝化螺旋菌能够在广泛的栖息地定居,并维持环境条件的变化,如氧气浓度的变化。
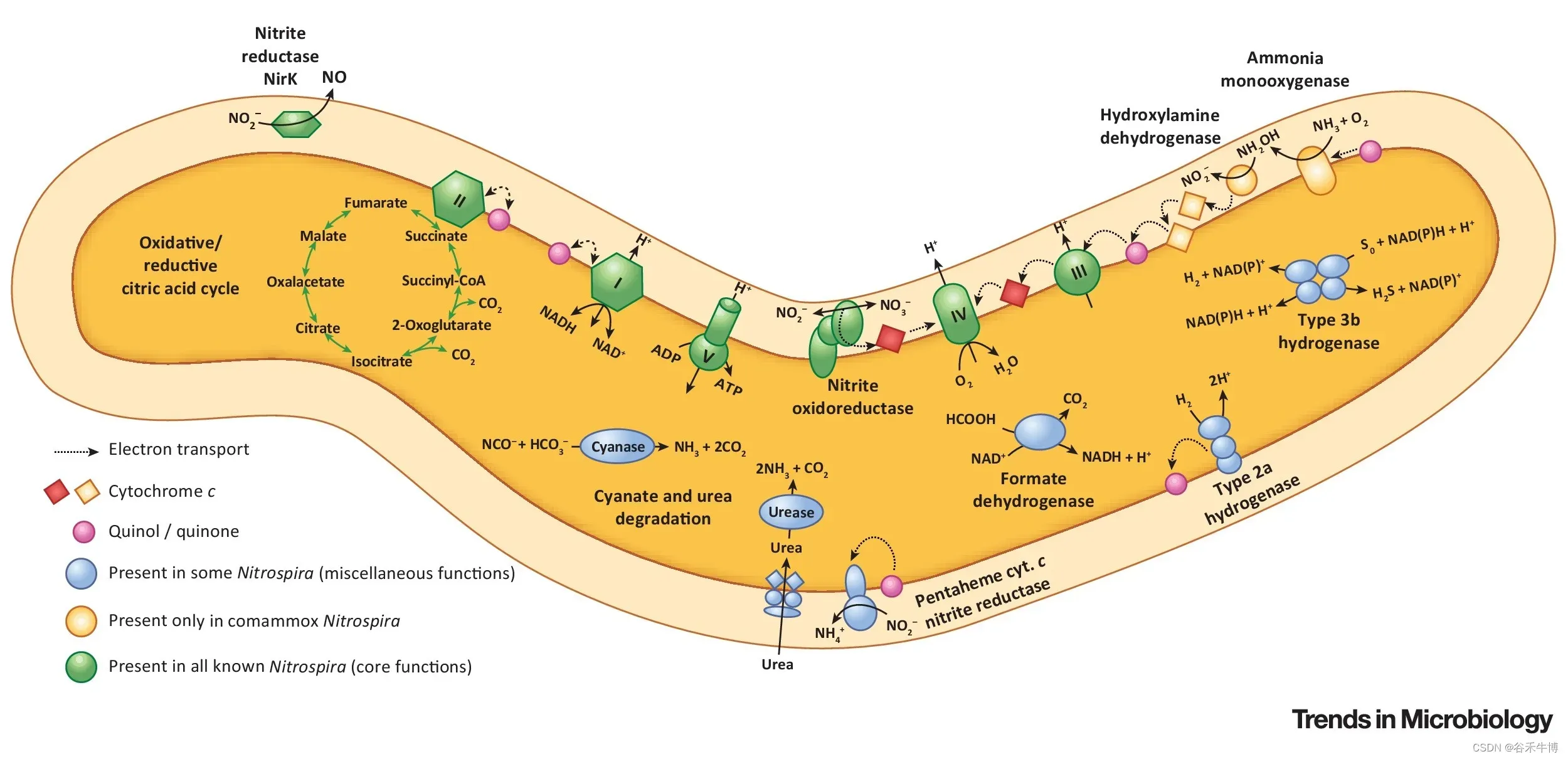
doi.org/10.1016/j.tim.2018.02.001
一些种类的硝化螺旋菌还能够利用硫化合物或铁等替代电子受体,进行厌氧呼吸。
已知一些种类的硝化螺旋菌参与有机物的降解,而另一些种类则参与甲烷的生产。
▼
健康相关
⇒
比较墨西哥城儿童(西方化,高动物蛋白和精制糖饮食)和 Me’phaa 儿童(非西方化,高纤维饮食)的肠道菌群,这两个人群主要区别在于不同种类的饮食。研究发现,Me’phaa 儿童表现出更高的绿弯菌门(Chloroflexi)和硝化螺旋菌(Nitrospirae)。
注:来自格雷罗州“Montaña Alta”地区的 Me’phaa 是一个前西班牙土著群体,他们主要靠种植豆类和扁豆,玉米等为生。还收集野生食用植物,并种植一些水果和蔬菜。肉类几乎只是在特殊场合食用的,并不是日常饮食的一部分。其生活方式和墨西哥城形成鲜明对比。
⇒
但也有研究发现,耐药癫痫患者表现出硝化螺旋菌富集(Kruskal-Wallis检验:p<0.05)。
⇒
一项关于胃微生物群的研究发现,硝螺旋菌门存在于所有胃癌患者中,但在慢性胃炎患者中完全不存在。
⇒
乳铁蛋白可以降低硝化螺旋菌水平。
No.12
脱 铁 杆 菌 门 Deferribacteres
脱铁杆菌(Deferribacteres)是1999年首次被描述的一门细菌。这些细菌以其还原铁和其他金属的能力而闻名,它们存在于各种环境中,包括深海热液喷口、温泉和地下水。
脱铁杆菌是革兰氏阴性细菌,通常是杆状的。它们是厌氧菌,通常在低氧环境中被发现。脱铁杆菌也是嗜热的,一些种类的脱铁杆菌能够进行化能生长,这意味着它们可以通过氧化无机化合物(如铁或硫)来获得能量。
G. thiophilus 阴性染色细胞的电子显微照

Janssen et al. 2002
脱铁杆菌中最著名的一种是脱铁杆菌脱硫菌,它能够还原铁和硫化合物,它被认为在深海环境中这些元素的循环中发挥着重要作用。
▼
代谢特性
脱铁杆菌属细菌最有趣的特征之一是它们能还原铁。这个过程包括将电子从铁转移到细菌,然后细菌可以利用这些电子产生能量。这一过程在许多环境中都很重要,因为它可以帮助维持生态系统中铁和其他金属的平衡。
除了还原铁,一些种类的脱铁杆菌还能还原其他金属,如锰和铀。脱铁杆菌属细菌在碳循环中也很重要,因为它们能够分解有机物并释放二氧化碳。一些种类的脱铁杆菌已知参与复杂有机化合物的降解,如木质素和纤维素。这一过程在许多环境中都很重要,因为它有助于循环营养物质,维持生态系统的平衡。
▼
健康相关
谷禾肠道数据库中大约4.92%的人群有检出。
⇒
脱铁杆菌门可能与环境高温高湿导致的肠道菌群失调和轻微肠炎有关。
研究分为三组实验:
- 对照组小鼠活跃,毛发光滑,排泄固体粪便。
- 高温高湿组中的小鼠都会出现粘性粪便,其中大约一半会分泌松散的粪便。
- 益生菌组经过益生菌治疗后,症状明显改善。
脱铁杆菌门是唯一在三组中具有差异丰度的门(P < 0.05),从正常对照组和高温高湿组的0.05%增加到广谱益生菌治疗组的1%。
⇒
一项研究发现,与正常对照和葡萄糖调节受损 (IGR) 患者相比,2型糖尿病患者中的脱铁杆菌门显着增加。
也发现脱铁杆菌门与镁摄入量呈负相关。
⇒
大鼠给予大麦或麦芽(7-8膳食纤维/100 g)4周,与对照大鼠相比,大麦组大鼠盲肠微生物群中的脱铁杆菌门的丰度低于对照组。
扩展阅读:谷物调节肠道菌群,促进代谢健康
在糖尿病中,关于脱铁杆菌门的研究不一致,有研究认为,糖尿病的改善与脱铁杆菌门相对丰度的降低有关,也有研究发现糖尿病的改善与脱铁杆菌门相对丰度的升高有关。
⇒
研究橄榄苦苷摄入对晚期2型糖尿病的缓解作用,发现橄榄苦苷可以增加Verrucomicrobia和脱铁杆菌门的相对丰度。
⇒
膳食菊粉治疗糖尿病组中脱铁杆菌门相对丰度下降。膳食菊粉通过抑制炎症和调节肠道微生物群来缓解2型糖尿病的不同阶段。
⇒
皮质酮治疗的小鼠中,拟杆菌门减少,脱铁杆菌门显著增加,水苏糖使拟杆菌门和脱铁杆菌恢复到正常水平。
⇒
较低的色氨酸补充量降低了脱铁杆菌门的丰度,而较高的色氨酸补充量不仅恢复了丰度,而且增加了丰度。
扩展阅读:色氨酸代谢与肠内外健康稳态
No.13
螺 旋 体 Spirochaetes
螺旋体(Spirochaetes),是革兰氏阴性菌,可运动的螺旋状细菌,断面呈圆形,以横向分裂繁殖,长度有 5 到 250 微米。螺旋体的独特之处在于,它们具有细胞内鞭毛。螺旋体属于双膜细菌门。

图片来源:crondon
螺旋体的细胞体被包裹在几层中。这些包括外膜和内膜、肽聚糖层以及细胞质膜。
其中一些是人类的严重病原体,会导致梅毒、雅司病、莱姆病和回归热等疾病。螺旋体属有螺旋体、密螺旋体、疏螺旋体、钩端螺旋体等。
▼
生存环境
螺旋体可以在水(地表水/淡水)、湖泊、盐沼沉积物、泥浆、沉积物、深海喷口、血液和淋巴等各种栖息地中找到。
图源:Oxford Nanopore Technologies
▼
代谢特性
螺旋体本质上是化学异养的,能够在厌氧条件下繁衍生息。
螺旋体的一个很好的例子是 Spirochaeta isovalerica。它是专性厌氧菌,它们通过发酵碳水化合物产生乙酸盐、乙醇、二氧化碳和氢气来生存。
许多种类的螺旋体对环境有益,并在固氮和有机物分解等过程中发挥重要作用。密螺旋体的某些种类生活在牛胃的瘤胃中,在那里它们为宿主分解纤维素和其他难以消化的植物多糖。
一些物种也被用于生物技术和工业应用,如生物燃料和生物塑料的生产。
▼
健康相关
谷禾肠道数据库中大约4.04%的人群有检出。
⇒
钩端螺旋菌病是由动物传染给人类的,一种常见的传播形式是让受污染的水接触到皮肤、眼睛和粘膜中未愈合的伤口。水由于与受感染动物的尿液接触而受到污染。——Leptospira
莱姆病 —— Borrelia burgdorferi,
Borrelia garinii, Borrelia afzelii
扩展阅读:夏季来临,警惕蜱虫叮咬感染疾病——莱姆病
回归热 ——复发性疏螺旋体(Borrelia recurrentis)
梅毒——梅毒螺旋体(Treponema pallidum)
雅司病(皮肤、骨骼和关节的热带感染)
—— T. pallidum subspeciespertenue
肠道螺旋体病——Brachyspira pirosicoli和Brachyspira aalborgi
文森特心绞痛—— Borrelia vincentii
⇒
在马来西亚北部的一个农村地区,与富裕人孩子相比,相对经济困难的土著儿童肠道微生物群表现出最多的微生物多样性。
Aeromonadales、拟杆菌门、瘤胃球菌科(Ruminococcaceae)、Deltaproteobacteria和螺旋体(Spirochaetes)富集,这与富含纤维的食物的分解有关。
No.14
装 甲 菌 门 Armatimonadetes
装甲菌门(Armatimonadetes),以前被称为候选门OP10,其成员分布在各种环境中,包括土壤、岩石、淡水和海洋沉积物。谷禾肠道数据库中大约3.31%的人群有检出。
装甲菌门细菌的特征是其独特的细胞壁结构,其中包含一层肽聚糖和一层类似于革兰氏阴性菌的外膜层。它们还具有形成长而有分支的细丝的能力。绿弯菌门是与装甲菌门亲缘关系最密切的正式门。
已经发现一些种类的装甲菌门在碳和氮循环等环境过程中发挥着重要作用。
在装甲菌门中,属于Armatimonasis纲的菌株 YO-36 T和属于Chthoonomonadetes纲的菌株 T49 T是唯一有效命名的分离株。
T49 T中枢代谢和碳固定通过常规糖酵解和三羧酸循环进行。T49 T的一个有趣特征是它在 4.7 到 5.8 的窄 pH 范围内生长。
▼
健康相关
很少有关于人体肠道菌群内装甲菌门的研究。
在一些关节炎患者的滑液中也检测到了Armatimonadetes门细菌,这些可能是游离污染物或机会性定植剂,而不是病原体。
在人体肠道中,装甲菌门的作用尚不清楚,待进一步研究和探索。
No.15
绿 菌 门 Chlorobi
绿菌门(Chlorobi)是一类光合细菌,也被称为绿菌门,谷禾肠道数据库中大约1.9%的人群有检出。
绿菌门是一类光合细菌,能够利用光能进行光合作用,产生能量和有机物质。
▼
常见菌属
在人体肠道菌群中,Chlorobi包含以下几个菌属和菌种:
- Chlorobium:
包括Chlorobium limicola、Chlorobium phaeobacteroides、Chlorobium tepidum等。
- Prosthecochloris:
包括Prosthecochloris aestuarii、Prosthecochloris vibrioformis等。
- Chloroherpeton:
包括Chloroherpeton thalassium等。
- Chloronema:
包括Chloronema giganteum等。
▼
健康相关
⇒
有研究显示B 族链球菌定植的孕妇肠道菌群中检测到大量的绿菌门(Chlorobi),同时还有大量Lentisphaerae、Parcubacteria、Chloroflexi、Gemmatimonadetes、Acidobacteria、Fusobacteria 、 Fibrobacteres。
GBS感染孕妇的OTU水平与炎症指标存在显着相关性。表明包括绿菌门在内的多种菌改变与 GBS 阳性孕妇的炎症状态和新生儿血气指标有关。
⇒
一项研究发现,酒精性肝病患者肠道菌群中,携带的厚壁菌门(p=0.03)和绿菌门(Chlorobi)(p=0.009)的数量存在显著差异。
同样,携带绿菌门(Chlorobi)(p=0.01)和coprothermobacterota(p=0.03)的患者的肝脏失代偿严重程度也有显著差异。
扩展阅读:深度解析 | 肠道菌群与慢性肝病,肝癌
很少有关于人体肠道菌群内绿菌门的研究。可能其存在来源于饮食摄入或环境。在人体肠道中,绿菌门的作用尚不清楚,它们的作用和功能尚待进一步研究和探索。
No.16
迷 踪 菌 门 Elusimicrobia
迷踪菌门(Elusimicrobia),也称为”隐微菌门”,谷禾肠道数据库中大约1.44%的人群有检出。
它们是一类非常小的细菌,通常直径只有0.2-0.4微米。迷踪菌门的细胞壁非常薄,甚至可以说是缺乏细胞壁,这使得它们对抗生素的抵抗力较弱。
▼
代谢特性
迷踪菌门中的一些菌种是共生菌,与其他生物共同生活,例如Candidatus Endomicrobium trichonymphae与白蚁肠道中的Trichonympha寄生在一起,共同分解木质素。
迷踪菌门的代谢功能多样,包括产生氢气、甲烷、酒精等。迷踪菌门在生态系统中扮演着重要的角色,例如在土壤中参与有机物分解、在海洋中参与碳循环等。
▼
健康相关
⇒
一项研究发现,在肥胖的2型糖尿病患者中,厚壁菌门丰度较高,而迷踪菌门(Elusimicrobia)丰度较低。
⇒
在两种大型急性辐射综合症动物模型的辐射暴露后的肠道菌群研究显示,哥廷根小型猪 (GMP)模型中的迷踪菌门在辐照后持续增加,表明它可用作肠道损伤的潜在生物标志物,以及对健康的潜在负面影响。
⇒
一项针对低质量睡眠和肠道菌群的研究显示Tenericutes 和 迷踪菌门(Elusimicrobia)在睡眠障碍患者中显著增加且与睡眠质量成正相关。
扩展阅读:肠道菌群与睡眠:双向调节
在人体肠道中,迷踪菌门的作用也还需要进一步研究。
No.17
衣 原 体 门 Chlamydiae
衣原体门(Chlamydiae),革兰氏阴性细菌,是专性寄生菌,它们的生长完全在其它生物的细胞内进行。
衣原体门细菌比一般细菌小,有的比病毒小,直径约为0.2-1.5微米。通常呈球形或椭圆形,没有细胞壁,但具有外膜和内膜。
编辑
图源:Science Photo Library
最开始,科学界普遍认为衣原体门的细胞壁不含肽聚糖,然而最近已有研究显示其细胞壁上的确有肽聚糖存在,并成功辨认出几种蛋白质。
衣原体是一类专性真核细胞内寄生、具有独特发育周期、可以在多种真核生物宿主(包括人、动物、原虫等)中繁殖的细菌,不能自主生长和繁殖。
▼
常见菌属
在人体肠道菌群中,Chlamydiae包含以下几个菌属和菌种:
- Chlamydia:
包括Chlamydia trachomatis、Chlamydia pneumoniae、Chlamydia psittaci 等。
- Parachlamydia:
包括Parachlamydia acanthamoebae、Parachlamydia boviseptica 等。
▼
传播方式
衣原体门是一种常见的病原体,可以引起多种疾病,包括性传播疾病、肺炎、结膜炎等。
衣原体的类型和相关疾病已知的与人类疾病有关的衣原体有三种,分别是鹦鹉热衣原体、沙眼衣原体和肺炎衣原体。这三种衣原体均可引起肺部感染。
鹦鹉热衣原体可通过感染有该种衣原体的禽类,如鹦鹉、孔雀、鸡、鸭、鸽等的组织、血液和粪便,以接触和吸入的方式感染给人类。
沙眼衣原体和肺炎衣原体主要在人类之间以呼吸道飞沫、母婴接触和性接触等方式传播。
▼
肠道健康相关
在谷禾肠道菌群数据库中大约有0.14%的人群有检出。
如果肠道感染衣原体,因感染肠道黏膜细胞可能导致腹泻、腹痛、恶心和呕吐,还可能会引发发热;腹泻和呕吐可能导致脱水、营养不良。
总之,衣原体作为一类常见的病原体,引起多种疾病,需要引起足够的重视和预防。
更多关于衣原体的介绍详见:
衣原体感染——原因、症状、治疗及预防
结 语
肠道微生物群一直伴随着人类的进化,并在人类的健康生活和高质量长寿中扮演着不可忽视的重要角色。
随着人类活动范围的不断扩大,一些以前未曾接触过的微生物会通过各种途径进入人体肠道,从而有机会在人体内生存和繁殖,并在人际之间传播。
同时,随着工业化的发展,我们原有的肠道菌群也面临着挑战,逐渐接受新的菌群、新的食物和添加剂等。
肠道菌群中一些占比较少的菌属,对人体健康和疾病的影响虽然研究较少,但这并不意味着它们的作用微弱。这些小众门派的菌属在自然界经过数亿年的进化选择,能够适应更恶劣的环境和在更寡营养条件下生存。此外,一些菌属具有特定的代谢功能,这些功能对我们的身体产生何种影响,以及它们如何影响人类的大尺度进化,都值得关注和研究。
尽管这些菌门的研究相对较少,但已经发现了一些有意思的结果。例如:
Akkermansia可以帮助减轻肥胖、改善胰岛素抵抗和代谢综合征等问题;具核梭杆菌(Fusobacterium nucleatum)与口腔疾病和肠炎结直肠癌的发病关系。莱姆病的罪魁祸首为伯氏疏螺旋体(Borrelia burgdorferi)。
在大人群水平上研究和探索这些菌属的来源及其代谢与人体健康的关系,将有助于发现和完善肠道菌群对人体健康的作用。这将为我们提供更好的方法来预防和治疗与肠道菌群相关的疾病,从而提高人类的健康水平。
主要参考文献:
Heinz Schlesner, Cheryl Jenkins,James T. Staley.The Phylum Verrucomicrobia: A Phylogenetically Heterogeneous Bacterial Group. The Prokaryotes pp 881–896
Hu C, Rzymski P. Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life (Basel). 2022 Mar 25;12(4):476. doi: 10.3390/life12040476. PMID: 35454968; PMCID: PMC9029806.
Verma D, Garg PK, Dubey AK. Insights into the human oral microbiome. Arch Microbiol. 2018 May;200(4):525-540. doi: 10.1007/s00203-018-1505-3. Epub 2018 Mar 23. PMID: 29572583.
Doocey CM, Finn K, Murphy C, Guinane CM. The impact of the human microbiome in tumorigenesis, cancer progression, and biotherapeutic development. BMC Microbiol. 2022 Feb 12;22(1):53. doi: 10.1186/s12866-022-02465-6. PMID: 35151278; PMCID: PMC8840051.
ohn D. Coates. (2010). Phylum XVII. Acidobacteria phyl. nov.
Miriam Gonçalves de Chaves. (2019). Acidobacteria Subgroups and Their Metabolic Potential for Carbon Degradation in Sugarcane Soil Amended With Vinasse and Nitrogen Fertilizers.
Sadaf Kalam et al. (2020). Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review.
Anna M. Kielak et al. (2016). The Ecology of Acidobacteria: Moving beyond Genes and Genomes.
Bong Suk Shim. (2011). Current Concepts in Bacterial Sexually Transmitted Diseases.
Jones and Bartlett Learning. Infectious Diseases Affecting the Respiratory System.
Yu-Jie Zhang, Sha Li, Ren-You Gan, and Tong Zhou. (2015). Impacts of Gut Bacteria on Human Health and Diseases.
Caitlin A. Brennan and Wendy S. Garrett. (2019). Fusobacterium nucleatum — symbiont, opportunist and oncobacterium.
K. W. Bennett and A. Eley. (1993). K. Fusobacteria: New taxonomy and related diseases Free.
Kevin Afra, Kevin Laupland, Jenine Leal, Tracie Lloyd, and Daniel Gregson. (2013). Incidence, risk factors, and outcomes of Fusobacterium species bacteremia.
Radhey Gupta and Mohit Sethi. (2014). Phylogeny and Molecular Signatures for the Phylum Fusobacteria and its Distinct Subclades.
John D. Coates. (2010). Phylum XVII. Acidobacteria phyl. nov.
Miriam Gonçalves de Chaves. (2019). Acidobacteria Subgroups and Their Metabolic Potential for Carbon Degradation in Sugarcane Soil Amended With Vinasse and Nitrogen Fertilizers.
Sadaf Kalam et al. (2020). Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review.
Anna M. Kielak et al. (2016). The Ecology of Acidobacteria: Moving beyond Genes and Genomes.
Zheng R, Liu R, Shan Y, Cai R, Liu G, Sun C. Characterization of the first cultured free-living representative of Candidatus Izemoplasma uncovers its unique biology. ISME J. 2021 Sep;15(9):2676-2691. doi: 10.1038/s41396-021-00961-7. Epub 2021 Mar 21. PMID: 33746205; PMCID: PMC8397711.
Bhandari V, Gupta RS. Molecular signatures for the phylum Synergistetes and some of its subclades. Antonie Van Leeuwenhoek. 2012 Nov;102(4):517-40. doi: 10.1007/s10482-012-9759-2. Epub 2012 Jun 19. PMID: 22711299.
Wang Z, Pu W, Liu Q, et al. Association of Gut Microbiota Composition in Pregnant Women Colonized with Group B Streptococcus with Maternal Blood Routine and Neonatal Blood-Gas Analysis. Pathogens. 2022;11(11):1297. Published 2022 Nov 4. doi:10.3390/pathogens11111297
He H, Lin M, You L, Chen T, Liang Z, Li D, Xie C, Xiao G, Ye P, Kong Y, Zhou Y. Gut Microbiota Profile in Adult Patients with Idiopathic Nephrotic Syndrome. Biomed Res Int. 2021 Feb 18;2021:8854969. doi: 10.1155/2021/8854969. PMID: 33681383; PMCID: PMC7910048.
Zhang J, Zhang X, Zhang K, et al. The Component and Functional Pathways of Gut Microbiota Are Altered in Populations with Poor Sleep Quality – A Preliminary Report. Pol J Microbiol. 2022;71(2):241-250. Published 2022 Jun 19. doi:10.33073/pjm-2022-021
Carbonero F, Mayta A, Bolea M, et al. Specific Members of the Gut Microbiota are Reliable Biomarkers of Irradiation Intensity and Lethality in Large Animal Models of Human Health. Radiat Res. 2019;191(1):107-121. doi:10.1667/RR14975.1
Lage OM, Bondoso J. Planctomycetes and macroalgae, a striking association. Front Microbiol. 2014 Jun 3;5:267. doi: 10.3389/fmicb.2014.00267. PMID: 24917860; PMCID: PMC4042473.
Kaboré OD, Godreuil S, Drancourt M. Planctomycetes as Host-Associated Bacteria: A Perspective That Holds Promise for Their Future Isolations, by Mimicking Their Native Environmental Niches in Clinical Microbiology Laboratories. Front Cell Infect Microbiol. 2020 Nov 30;10:519301. doi: 10.3389/fcimb.2020.519301. PMID: 33330115; PMCID: PMC7734314.
Bell A, Severi E, Owen CD, Latousakis D, Juge N. Biochemical and structural basis of sialic acid utilization by gut microbes. J Biol Chem. 2023 Feb 8;299(3):102989. doi: 10.1016/j.jbc.2023.102989. Epub ahead of print. PMID: 36758803; PMCID: PMC10017367.
Wang J, Wang Y, Gao W, Wang B, Zhao H, Zeng Y, Ji Y, Hao D. Diversity analysis of gut microbiota in osteoporosis and osteopenia patients. PeerJ. 2017 Jun 15;5:e3450. doi: 10.7717/peerj.3450. PMID: 28630804; PMCID: PMC5474093.
Bakhti SZ, Latifi-Navid S. Interplay and cooperation of Helicobacter pylori and gut microbiota in gastric carcinogenesis. BMC Microbiol. 2021 Sep 23;21(1):258. doi: 10.1186/s12866-021-02315-x. PMID: 34556055; PMCID: PMC8461988.
Glover JS, Ticer TD, Engevik MA. Characterizing the mucin-degrading capacity of the human gut microbiota. Sci Rep. 2022 May 19;12(1):8456. doi: 10.1038/s41598-022-11819-z. PMID: 35589783; PMCID: PMC9120202.
Wu N, Zhou J, Mo H, Mu Q, Su H, Li M, Yu Y, Liu A, Zhang Q, Xu J, Yu W, Liu P, Liu G. The Gut Microbial Signature of Gestational Diabetes Mellitus and the Association With Diet Intervention. Front Cell Infect Microbiol. 2022 Jan 14;11:800865. doi: 10.3389/fcimb.2021.800865. PMID: 35096649; PMCID: PMC8795975.
Castillo-Álvarez F, Pérez-Matute P, Oteo JA, Marzo-Sola ME. The influence of interferon β-1b on gut microbiota composition in patients with multiple sclerosis. Neurologia (Engl Ed). 2021 Sep;36(7):495-503. doi: 10.1016/j.nrleng.2020.05.006. Epub 2020 May 31. PMID: 34537163.
Zhang Y, Lu S, Yang Y, Wang Z, Wang B, Zhang B, Yu J, Lu W, Pan M, Zhao J, Guo S, Cheng J, Chen X, Hong K, Li G, Yu Z. The diversity of gut microbiota in type 2 diabetes with or without cognitive impairment. Aging Clin Exp Res. 2021 Mar;33(3):589-601. doi: 10.1007/s40520-020-01553-9. Epub 2020 Apr 16. PMID: 32301029.
Harakeh S, Angelakis E, Karamitros T, Bachar D, Bahijri S, Ajabnoor G, Alfadul SM, Farraj SA, Al Amri T, Al-Hejin A, Ahmed A, Mirza AA, Didier R, Azhar EI. Impact of smoking cessation, coffee and bread consumption on the intestinal microbial composition among Saudis: A cross-sectional study. PLoS One. 2020 Apr 29;15(4):e0230895. doi: 10.1371/journal.pone.0230895. PMID: 32348307; PMCID: PMC7190147.
Fuerst JA, Sagulenko E. Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function. Nat Rev Microbiol. 2011 Jun;9(6):403-13. doi: 10.1038/nrmicro2578. PMID: 21572457.
Gong X, Liu X, Chen C, Lin J, Li A, Guo K, An D, Zhou D, Hong Z. Alteration of Gut Microbiota in Patients With Epilepsy and the Potential Index as a Biomarker. Front Microbiol. 2020 Sep 18;11:517797. doi: 10.3389/fmicb.2020.517797. PMID: 33042045; PMCID: PMC7530173.
Yuan X, Chen R, McCormick KL, Zhang Y, Lin X, Yang X. The role of the gut microbiota on the metabolic status of obese children. Microb Cell Fact. 2021 Feb 27;20(1):53. doi: 10.1186/s12934-021-01548-9. PMID: 33639944; PMCID: PMC7916301..
Zhuang X, Tian Z, Li L, Zeng Z, Chen M, Xiong L. Fecal Microbiota Alterations Associated With Diarrhea-Predominant Irritable Bowel Syndrome. Front Microbiol. 2018 Jul 25;9:1600. doi: 10.3389/fmicb.2018.01600. PMID: 30090090; PMCID: PMC6068233.
Polo PG, Çolak-Al B, Sentürk H, Rafiqi AM. Gut bacteria after recovery from COVID-19: a pilot study. Eur Rev Med Pharmacol Sci. 2022 Nov;26(22):8599-8611. doi: 10.26355/eurrev_202211_30397. PMID: 36459041.
Dix C, Wright O. Bioavailability of a Novel Form of Microencapsulated Bovine Lactoferrin and Its Effect on Inflammatory Markers and the Gut Microbiome: A Pilot Study. Nutrients. 2018 Aug 17;10(8):1115. doi: 10.3390/nu10081115. PMID: 30126153; PMCID: PMC6115941.
Sánchez-Quinto A, Cerqueda-García D, Falcón LI, Gaona O, Martínez-Correa S, Nieto J, G-Santoyo I. Gut Microbiome in Children from Indigenous and Urban Communities in México: Different Subsistence Models, Different Microbiomes. Microorganisms. 2020 Oct 16;8(10):1592. doi: 10.3390/microorganisms8101592. PMID: 33081076; PMCID: PMC7602701.
Stewart OA, Wu F, Chen Y. The role of gastric microbiota in gastric cancer. Gut Microbes. 2020 Sep 2;11(5):1220-1230. doi: 10.1080/19490976.2020.1762520. Epub 2020 May 23. PMID: 32449430; PMCID: PMC7524314.
Chen S, Zheng Y, Zhou Y, Guo W, Tang Q, Rong G, Hu W, Tang J, Luo H. Gut Dysbiosis with Minimal Enteritis Induced by High Temperature and Humidity. Sci Rep. 2019 Dec 10;9(1):18686. doi: 10.1038/s41598-019-55337-x. PMID: 31822775; PMCID: PMC6904617.
Do MH, Lee HB, Oh MJ, Jhun H, Choi SY, Park HY. Polysaccharide fraction from greens of Raphanus sativus alleviates high fat diet-induced obesity. Food Chem. 2021 May 1;343:128395. doi: 10.1016/j.foodchem.2020.128395. Epub 2020 Oct 15. PMID: 33268179.
Phillippi DT, Daniel S, Pusadkar V, Youngblood VL, Nguyen KN, Azad RK, McFarlin BK, Lund AK. Inhaled diesel exhaust particles result in microbiome-related systemic inflammation and altered cardiovascular disease biomarkers in C57Bl/6 male mice. Part Fibre Toxicol. 2022 Feb 9;19(1):10. doi: 10.1186/s12989-022-00452-3. PMID: 35135577; PMCID: PMC8827295.
Nuli R, Cai J, Kadeer A, Zhang Y, Mohemaiti P. Integrative Analysis Toward Different Glucose Tolerance-Related Gut Microbiota and Diet. Front Endocrinol (Lausanne). 2019 May 27;10:295. doi: 10.3389/fendo.2019.00295. PMID: 31191448; PMCID: PMC6546033.
Zhong Y, Nyman M, Fåk F. Modulation of gut microbiota in rats fed high-fat diets by processing whole-grain barley to barley malt. Mol Nutr Food Res. 2015 Oct;59(10):2066-76. doi: 10.1002/mnfr.201500187. Epub 2015 Aug 26. PMID: 26184884.
Shafiee NH, Razalli NH, Muhammad Nawawi KN, Mohd Mokhtar N, Raja Ali RA. Implication of food insecurity on the gut microbiota and its potential relevance to a multi-ethnic population in Malaysia. JGH Open. 2022 Feb 1;6(2):112-119. doi: 10.1002/jgh3.12709. PMID: 35155820; PMCID: PMC8829104.
Lee KC, Herbold CW, Dunfield PF, Morgan XC, McDonald IR, Stott MB. Phylogenetic delineation of the novel phylum Armatimonadetes (former candidate division OP10) and definition of two novel candidate divisions. Appl Environ Microbiol. 2013 Apr;79(7):2484-7. doi: 10.1128/AEM.03333-12. Epub 2013 Feb 1. PMID: 23377935; PMCID: PMC3623213.
版权声明:本文为博主作者:谷禾牛博原创文章,版权归属原作者,如果侵权,请联系我们删除!
原文链接:https://blog.csdn.net/Hangzhou_Guhe/article/details/129793370